■DNAコンピュータ
- DNAの基礎知識
DNAコンピュータは生体、とくにDNAの複製の仕組みを真似ることに端を発している。そのためDNAコンピュータを理解するには、分子生物学の基本的な知識が必要となる。このページでは、DNAコンピュータと関わりの深いものについて簡単に触れておく。
DNAの基本構造と複製機能
遺伝情報が親から子へと受け継がれていくためには、親とまったく同じDNAが合成される必要がある。この正確なDNAの複製には、たった4つの塩基対が大きな役割を果たしている。アデニン(A)、グアニン(G)、チミン(T)、シトシン(C)の塩基対は、塩基対どうしの水素結合によって、AとT,CとGという選択的なペアを形成している。DNAが複製されるためには、いったんこの水素結合をたち切って、DNAの二重らせんをほどいてやる必要がある。いったん二重らせん構造がほどかれると、一本鎖のDNAはさまざまな酵素のはたすけをかりて巧みに複製を行なっていく。その様子は次のようなものだ。
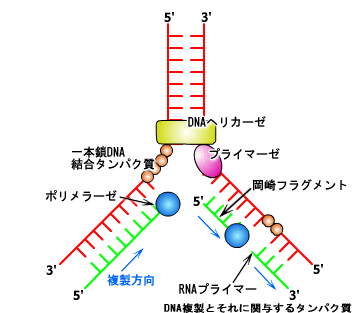
二重らせん構造をほどかれて一本鎖になったDNAに、「DNAポリメラーゼ」と呼ばれる酵素がとりつき、まるでレールの上を滑っていくかのように、重合を起こして鋳型となったDNAと相補的なDNAをあらたに合成する。ただし、DNAポリメラーゼが鋳型のDNAを進んでいく方向には決まりがあり、5'→3'方向へしか進むことができない。
ところが二重らせん構造をとっているDNAは、一方は5'→3'だが、もう片方は必ず3'→5'になっている。しかし、DNAポリメラーゼはこの方向に伸長を触媒することはできない。そのためDNAポリメラーゼは、5'→3'のときのように連続的にではなく、図のように100~1000程度の短いDNAを不連続に新しいDNAを合成している。この断片的なDNAの構造を発見者にちなんで岡崎フラグメントと呼んでいる。この断片は「DNAリガーゼ」と呼ばれる酵素によって糊づけされるかのように連結されて長いDNA鎖になる。複製方向に一致する側に連続的に合成される娘鎖をリーディング鎖、複製方向に逆行し合成が不連続になる側の娘鎖をラギング鎖と呼んでいる。
実際のDNAの複製にはさらに多くの酵素やタンパク質が関与している。例えば、二重らせんをほどいて一本鎖をまっすぐにしているのは「DNAヘリカーゼ」だ。また、ラギング鎖でDNAポリメラーゼが合成を開始するきっかけをつくるのが、「プライマー」と呼ばれる、通常3~5個程度の塩基である。このプライマーを合成しているのが「プライマーゼ」と呼ばれる酵素である。
DNAの増幅 - ポリメラーゼ連鎖反応(PCR)
しかしその出力結果を得るとき、一つ一つのDNA分子だけで認識することは難しい。実際にはゲル電気泳動(下参考)などによって、特定のDNA分子の集団を認識しているので、同じ塩基配列を持ったDNAを大量に用意する必要がある。その方法が「ポリメラーゼ連鎖反応(PCR;Polymerase Chain Reaction)」と呼ばれる技術である。
具体的にPCRは次のような方法で行なわれる。
①増幅させたいDNAを94℃で加熱して一本鎖にする。
②それぞれの鎖に相補的なプライマーをそれぞれ55~60℃程度の環境下で固定する。
③DNAポリメラーゼを用いて72℃でDNAの合成を行なう。
④合成されたDNAを再度一本かしてDNA合成反応を繰り返す。
こうすることでn回のサイクルにより、理論上は目的のDNA鎖が2^n個(実際は2^n-2n,ただしnが大きいときには2^nと見なしてかまわない)できることになる。だいたい三時間ほどで20~50万倍ほどに増幅できる。現在PCRは実に多様な方面で重要な利用法が考え出されている。例えば、ウィルスや細菌の検出、遺伝子クローニング、遺伝子変異導入などがそうである。なお、はじめにPCRを考案したK.B.Mullisらは、この業績をたたえられ1993年にノーベル化学賞を受けている。
ゲル電気泳動
複数種のDNA分子が混ざった溶液を寒天のようなゲル状の板の一端に置いて、板に電気を流す。すると、DNA分子は負の電荷をもっているので陰極へと移動するが、このとき分子量の小さいものは大きいものより速く移動する。したがって、DNAの分子量、つまり長さによって分離することができる。特殊な化合物を使って紫外線を照射することで、DNAの移動後の様子が縞模様(バンド)としてみることができる。
制限酵素
「制限酵素(restriction enzyme)」は、DNA一本鎖上で特定の塩基配列を認識し、そのDNAを切断するという役割をもっている。制限酵素はDNAリガーゼとともに、60年代以降の組換えDNA技術で、非常に重要な位置を占めてきた。制限酵素で有名なものには、例えば"EcoRI"がある。

制限酵素"EcoRI"とDNAリガーゼによる組換えDNAの作製 |
この酵素は、"GAATTC"という塩基配列を認識し、GとAの間で図のように切断する。すると、片割れのDNA断片の部分が生じるが、この箇所は水素結合で他のDNA断片と結合しやすいため、粘着末端(sticky
end)と呼ばれている。この粘着末端を利用して、他の生物のDNAをDNAリガーゼを利用して接合することで組換えDNAができあがる。
この制限酵素EcoRIは大腸菌(Escherichia coli)から発見された酵素であるが、生体にとっての制限酵素のおもな役割は、バクテリアのような宿主組織をウィルスのような侵略者から守ることである。つまり侵略者のDNAを分解してしまうというものだ。ただし、このときに宿主組織のDNAも分解されてしまっては意味がないので、これを防ぐためにメチラーゼと呼ばれる「修飾酵素(modifying enzyme)」で特定のヌクレオチドをメチル化して制限酵素がDNAにとりつけないようにしている。そのため修飾酵素は、よく制限酵素とペアとして考えられる。
DNAの分子演算
ここのページで紹介したいくつかのDNA関連の酵素は、DNAコンピューティングに必要な演算子としての性質をもっている。次のページ以降で具体的な方法について述べるが、ここで簡単にまとめておこう。これらの演算はDNAコンピューティングのほとんどのアルゴリズムにおいて基本的に共通している。
| DNAの分子演算 |
説明 |
| 融解(melting) |
ある特定の温度で溶液を熱することにより、二本鎖のDNAが一本鎖に分離するこのように相補的分子間の水素結合が壊れる。 |
| アニ-リング(annealing) |
これは融解の逆演算である。一本鎖分子の溶液が冷やされると、ワトソン・クリック相補的な分子どうしが互いに結合する。 |
| マージ(merge) |
二つの試験管の内容物を一本の試験管に流し込んで一緒にする。 |
| 増幅(amplification) |
これはポリメラーゼ連鎖反応(PCR)を適用することに他ならない。 |
| 検知と長さによる分離 |
ゲル電気泳動 |
なお、すべてのDNAコンピューティングに共通しているとはいえないが、制限酵素などを使ったスプライシング(切断)演算も、複雑なDNAコンピューティングを実現するには重要である。これについては、「DNAコンピューティング/さまざまな酵素を使った複雑なDNAコンピュータ」で具体例を紹介している。
|
|